
Semua konten iLive ditinjau secara medis atau diperiksa fakta untuk memastikan akurasi faktual sebanyak mungkin.
Kami memiliki panduan sumber yang ketat dan hanya menautkan ke situs media terkemuka, lembaga penelitian akademik, dan, jika mungkin, studi yang ditinjau secara medis oleh rekan sejawat. Perhatikan bahwa angka dalam tanda kurung ([1], [2], dll.) Adalah tautan yang dapat diklik untuk studi ini.
Jika Anda merasa salah satu konten kami tidak akurat, ketinggalan zaman, atau dipertanyakan, pilih dan tekan Ctrl + Enter.
Ovarium
Ahli medis artikel

Terakhir ditinjau: 04.07.2025

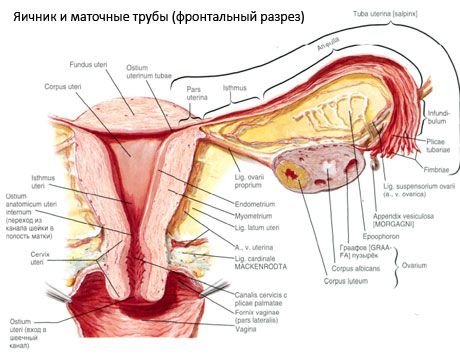
Ovarium (ovarium; ooforon Yunani) adalah organ berpasangan, kelenjar reproduksi wanita, yang terletak di rongga panggul di belakang ligamen uterus yang luas. Di ovarium, sel-sel reproduksi wanita (telur) berkembang dan matang, dan hormon seks wanita terbentuk yang memasuki darah dan getah bening. Ovarium memiliki bentuk lonjong, agak pipih di arah anteroposterior. Warna ovarium merah muda. Di permukaan ovarium wanita yang telah melahirkan, depresi dan bekas luka terlihat - jejak ovulasi dan transformasi korpora lutea. Ovarium beratnya 5-8 g. Dimensi ovarium adalah: panjang 2,5-5,5 cm, lebar 1,5-3,0 cm, tebal - hingga 2 cm. Ovarium memiliki dua permukaan bebas: permukaan medial (facies medialis), menghadap rongga panggul, sebagian tertutup oleh tuba fallopi, dan permukaan lateral (facies lateralis), berdekatan dengan dinding samping panggul, hingga lekukan yang sedikit menonjol - fossa ovarium. Fossa ini terletak di sudut antara pembuluh iliaka eksternal yang ditutupi oleh peritoneum di bagian atas dan arteri uterus dan obturator di bagian bawah. Di belakang ovarium, ureter dari sisi yang sesuai melewati retroperitoneal dari atas ke bawah.
Permukaan ovarium masuk ke tepi bebas cembung (posterior) (margo liber), di depan - ke tepi mesenterika (margo mesovaricus), dilekatkan melalui lipatan pendek peritoneum (mesenterium ovarium) ke selebaran posterior ligamentum latum uterus. Di tepi anterior organ ini terdapat lekukan beralur - hilum ovarium (hilum ovarii), tempat arteri dan saraf memasuki ovarium, dan vena serta pembuluh limfatik keluar. Ovarium juga memiliki dua ujung: ujung tubular atas yang membulat (extremitas tubaria), menghadap tuba falopi, dan ujung uterus bawah (extremitas utenna), yang dihubungkan ke uterus oleh ligamentum ovarium (lig. ovarii proprium). Ligamentum ini, dalam bentuk tali bundar setebal sekitar 6 mm, membentang dari ujung uterus ovarium ke sudut lateral uterus, yang terletak di antara dua lembar ligamentum latum uterus. Aparatus ligamentum ovarium juga mencakup ligamentum suspensori ovarium (lig.suspensorium ovarii), yang merupakan lipatan peritoneum yang membentang dari dinding pelvis minor ke ovarium dan berisi pembuluh ovarium dan berkas serat fibrosa di dalamnya. Ovarium difiksasi oleh mesenterium pendek (mesovarium), yang merupakan duplikasi peritoneum yang membentang dari daun posterior ligamentum latum uterus ke tepi mesenterika ovarium. Ovarium sendiri tidak ditutupi oleh peritoneum. Pinggiran ovarium terbesar dari tuba fallopi melekat pada ujung tubular ovarium. Topografi ovarium bergantung pada posisi uterus, ukurannya (selama kehamilan). Ovarium adalah organ yang sangat mobile dari rongga pelvis minor.
 [
[ Pembuluh darah dan saraf ovarium
Suplai darah ke ovarium disediakan oleh aa. et vv. ovaricae et uterinae. Kedua arteri ovarium (aa. ovaricae dextra et sinistra) berasal dari permukaan anterior aorta tepat di bawah arteri renalis; yang kanan biasanya berasal dari aorta, dan yang kiri dari arteri renalis. Mengarah ke bawah dan lateral sepanjang permukaan anterior otot psoas mayor, setiap arteri ovarium melintasi ureter di depan (memberikan cabang kepadanya), pembuluh iliaka eksternal, garis batas dan memasuki rongga panggul, yang terletak di sini di ligamen suspensorium ovarium. Mengikuti arah medial, arteri ovarium melewati antara daun ligamen latum uterus di bawah tuba falopi, memberikan cabang kepadanya, dan kemudian ke mesenterium ovarium; ia memasuki hilum ovarium.
Cabang-cabang arteri ovarium beranastomosis luas dengan cabang-cabang ovarium dari arteri uterus. Aliran keluar vena dari ovarium dilakukan terutama ke pleksus vena ovarium, yang terletak di daerah hilus ovarium. Dari sini, aliran keluar darah melewati dua arah: melalui vena uterus dan ovarium. Vena ovarium kanan memiliki katup dan mengalir ke vena cava inferior. Vena ovarium kiri mengalir ke vena renalis kiri, meskipun tidak memiliki katup.
Drainase limfatik dari ovarium terjadi melalui pembuluh limfatik, terutama di area gerbang organ, tempat pleksus limfatik subovarium dibedakan. Kemudian limfa dikeluarkan melalui pembuluh limfatik ovarium ke kelenjar getah bening paraaorta.
Persarafan ovarium
Simpatik - disediakan oleh serabut postganglionik dari pleksus seliaka (solar), mesenterika superior, dan hipogastrikus; parasimpatis - oleh saraf sakral viseral.
Struktur ovarium
Permukaan ovarium ditutupi dengan epitel germinal satu lapis. Di bawahnya terdapat lapisan protein jaringan ikat padat (tunika albuginea). Jaringan ikat ovarium membentuk stromanya (stroma ovarii), yang kaya akan serat elastis. Substansi ovarium, parenkimnya, terbagi menjadi lapisan luar dan dalam. Lapisan dalam, yang terletak di tengah ovarium, lebih dekat ke gerbangnya, disebut medula (medulla ovarii). Di lapisan ini, dalam jaringan ikat longgar, terdapat banyak pembuluh darah dan limfatik serta saraf. Lapisan luar ovarium - korteks (korteks ovarii) lebih padat. Ini mengandung banyak jaringan ikat, di mana terdapat folikel ovarium primer yang matang (folliculi ovarici primarii), folikel sekunder (vesikular) (folliculi ovarici secundarii, s.vesiculosi), serta folikel matang, folikel Graaf (folliculi ovarici maturis), serta badan kuning dan atretik.
Setiap folikel mengandung sel telur reproduksi betina, atau oosit (ovocytus). Sel telur berdiameter hingga 150 µm, berbentuk bulat, mengandung nukleus, sejumlah besar sitoplasma, yang, selain organel sel, mengandung inklusi protein-lipid (kuning telur), glikogen, yang diperlukan untuk nutrisi sel telur. Sel telur biasanya menghabiskan pasokan nutrisinya dalam waktu 12-24 jam setelah ovulasi. Jika pembuahan tidak terjadi, sel telur akan mati.
Sel telur manusia memiliki dua membran yang melapisinya. Di dalamnya terdapat sitolema, yang merupakan membran sitoplasma sel telur. Di luar sitolema terdapat lapisan yang disebut sel folikel, yang melindungi sel telur dan menjalankan fungsi pembentukan hormon - yaitu mengeluarkan estrogen.
Posisi fisiologis uterus, tuba, dan ovarium disediakan oleh alat-alat yang menahan, memperbaiki, dan menyokong yang menyatukan peritoneum, ligamen, dan jaringan panggul. Alat-alat yang menahan diwakili oleh formasi berpasangan, yang meliputi ligamen bundar dan lebar uterus, ligamen yang tepat, dan ligamen yang menahan ovarium. Ligamen yang lebar uterus, ligamen yang tepat, dan ligamen yang menahan ovarium menahan uterus di posisi tengah. Ligamen bundar menarik fundus uterus ke depan dan memberikan kemiringan fisiologisnya.
Alat pengikat (penahan) memastikan posisi rahim di tengah panggul kecil dan membuatnya hampir tidak mungkin bergeser ke samping, maju mundur. Namun karena alat ligamen berangkat dari rahim di bagian bawahnya, kemiringan rahim ke berbagai arah dimungkinkan. Alat pengikat meliputi ligamen yang terletak di jaringan longgar panggul dan memanjang dari bagian bawah rahim ke dinding lateral, anterior, dan posterior panggul: ligamen sakro-magik, kardinal, uterovesika, dan vesikopubik.
Selain mesovarium, ligamen ovarium berikut dibedakan:
- ligamentum suspensori ovarium, yang sebelumnya disebut ligamentum infundibulopelvikum. Ligamentum ini merupakan lipatan peritoneum dengan pembuluh darah (a. et v. ovarica) dan pembuluh limfatik serta saraf ovarium yang melewatinya, membentang di antara dinding lateral pelvis, fasia lumbal (di area pembagian arteri iliaka komunis menjadi eksternal dan internal) dan ujung atas (tuba) ovarium;
- ligamentum ovarium yang tepat melewati antara daun ligamentum uterus yang lebar, lebih dekat ke daun posterior, dan menghubungkan ujung bawah ovarium dengan tepi lateral uterus. Ligamentum ovarium yang tepat melekat pada uterus antara awal tuba fallopi dan ligamentum rotundum, di belakang dan di atas yang terakhir. Dalam ketebalan ligamentum tersebut melewati rr. ovarii, yang merupakan cabang terminal dari arteri uterus;
- ligamentum apendikular-ovarium Ligamentum ini membentang dari mesenterium apendiks ke ovarium kanan atau ligamentum latum uterus dalam bentuk lipatan peritoneum. Ligamentum ini tidak permanen dan ditemukan pada 1/2 - 1/3 wanita.
Peralatan pendukung diwakili oleh otot dan fasia dasar panggul, dibagi menjadi lapisan bawah, tengah, dan atas (dalam).
Lapisan otot yang paling kuat adalah lapisan otot atas (dalam), yang diwakili oleh otot berpasangan yang mengangkat anus. Lapisan ini terdiri dari kumpulan otot yang menyebar dari tulang ekor ke tulang panggul dalam tiga arah (pubococcygeus, iliococcygeus, dan ischiococcygeus). Lapisan otot ini juga disebut diafragma panggul.
Lapisan otot tengah terletak di antara simfisis, tulang kemaluan, dan tulang iskia. Lapisan otot tengah - diafragma urogenital - menempati separuh anterior pintu keluar panggul, tempat uretra dan vagina lewat. Di bagian anterior di antara lembarannya terdapat kumpulan otot yang membentuk sfingter eksternal uretra, di bagian posterior terdapat kumpulan otot yang mengarah ke arah melintang - otot transversal dalam perineum.
Lapisan bawah (luar) otot dasar panggul terdiri dari otot-otot superfisial yang bentuknya menyerupai angka 8. Otot-otot tersebut meliputi otot bulbokavernosus, otot iskiokavernosa, otot sfingter ani eksterna, dan otot perineum transversal superfisial.
Ontogenesis ovarium
Proses pertumbuhan folikel dan atresia dimulai pada minggu ke-20 kehamilan, dan pada saat melahirkan, hingga 2 juta oosit tetap berada di ovarium gadis itu. Pada menarche, jumlahnya berkurang menjadi 300 ribu. Selama seluruh periode kehidupan reproduksi, tidak lebih dari 500 folikel mencapai kematangan dan berovulasi. Pertumbuhan awal folikel tidak bergantung pada stimulasi FSH, terbatas, dan atresia cepat terjadi. Dipercayai bahwa alih-alih hormon steroid, peptida autokrin/parakrin lokal adalah pengatur utama pertumbuhan dan atresia folikel primer. Dipercayai bahwa proses pertumbuhan folikel dan atresia tidak terganggu oleh proses fisiologis apa pun. Proses ini berlanjut pada semua usia, termasuk periode intrauterin dan menopause, dan terganggu oleh kehamilan, ovulasi, dan anovulasi. Mekanisme yang memicu pertumbuhan folikel dan jumlahnya dalam setiap siklus spesifik masih belum jelas.
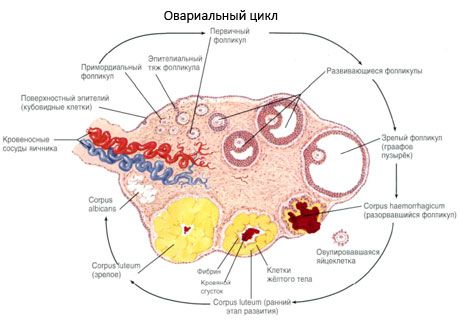
Selama perkembangannya, folikel mengalami beberapa tahap perkembangan. Sel germinal primordial berasal dari endoderm kantung kuning telur, allantois, dan bermigrasi ke area genital embrio pada usia kehamilan 5-6 minggu. Sebagai hasil dari pembelahan mitosis cepat, yang berlanjut dari usia kehamilan 6-8 minggu hingga 16-20 minggu, hingga 6-7 juta oosit terbentuk di ovarium embrio, dikelilingi oleh lapisan tipis sel granulosa.
Folikel preantral - oosit dikelilingi oleh membran (Zona pelusida). Sel-sel granulosa yang mengelilingi oosit mulai berkembang biak, pertumbuhannya bergantung pada gonadotropin dan berkorelasi dengan kadar estrogen. Sel-sel granulosa merupakan target FSH. Pada tahap folikel preantral, sel-sel granulosa mampu mensintesis tiga kelas steroid: terutama menginduksi aktivitas aromatase, enzim utama yang mengubah androgen menjadi estradiol. Dipercayai bahwa estradiol mampu meningkatkan jumlah reseptornya sendiri, memberikan efek mitogenik langsung pada sel-sel granulosa terlepas dari FSH. Ini dianggap sebagai faktor parakrin yang meningkatkan efek FSH, termasuk aktivasi proses aromatisasi.
Reseptor FSH muncul pada membran sel granulosa segera setelah pertumbuhan folikel dimulai. Penurunan atau peningkatan FSH menyebabkan perubahan jumlah reseptornya. Tindakan FSH ini dimodulasi oleh faktor pertumbuhan. FSH bekerja melalui protein G, sistem adenilat siklase, meskipun steroidogenesis dalam folikel sebagian besar diatur oleh FSH, banyak faktor yang terlibat dalam proses ini: saluran ion, reseptor tirosin kinase, sistem fosfolipase pembawa pesan kedua.
Peran androgen dalam perkembangan folikel dini sangatlah kompleks. Sel granulosa memiliki reseptor androgen. Reseptor ini tidak hanya merupakan substrat untuk aromatisasi yang diinduksi FSH menjadi estrogen, tetapi juga dapat meningkatkan aromatisasi pada konsentrasi rendah. Ketika kadar androgen meningkat, sel granulosa preantral lebih memilih jalur aromatisasi menjadi estrogen daripada jalur androgen yang lebih sederhana melalui 5a-reduktase untuk diubah menjadi androgen yang tidak dapat diubah menjadi estrogen, sehingga menghambat aktivitas aromatase. Proses ini juga menghambat pembentukan reseptor FSH dan LH, sehingga menghentikan perkembangan folikel.
Proses aromatisasi, folikel dengan kadar androgen tinggi mengalami proses atresia. Pertumbuhan dan perkembangan folikel bergantung pada kemampuannya mengubah androgen menjadi estrogen.
Dengan adanya FSH, substansi dominan dari cairan folikel adalah estrogen. Dengan tidak adanya FSH - androgen. LH biasanya tidak ada dalam cairan folikel sampai pertengahan siklus. Begitu kadar LH meningkat, aktivitas mitosis sel granulosa menurun, perubahan degeneratif muncul dan kadar androgen dalam folikel meningkat. Kadar steroid dalam cairan folikel lebih tinggi daripada dalam plasma dan mencerminkan aktivitas fungsional sel ovarium: sel granulosa dan sel teka. Jika satu-satunya target untuk FSH adalah sel granulosa, maka LH memiliki banyak target - sel teka, sel stroma dan lutein dan sel granulosa. Baik sel granulosa maupun sel teka memiliki kemampuan untuk steroidogenesis, tetapi aktivitas aromatase mendominasi dalam sel granulosa.
Sebagai respons terhadap LH, sel teka menghasilkan androgen, yang kemudian diubah oleh sel granulosa menjadi estrogen melalui aromatisasi yang diinduksi FSH.
Saat folikel tumbuh, sel-sel teka mulai mengekspresikan gen untuk reseptor LH, P450 sec dan 3beta-hydroxysteroid dehidrogenase, faktor pertumbuhan mirip insulin (IGF-1) secara sinergis dengan LH meningkatkan ekspresi gen, tetapi tidak merangsang steroidogenesis.
Steroidogenesis ovarium selalu bergantung pada LH. Saat folikel tumbuh, sel-sel teka mengekspresikan enzim P450c17, yang membentuk androgen dari kolesterol. Sel-sel granulosa tidak memiliki enzim ini dan bergantung pada sel-sel teka untuk menghasilkan estrogen dari androgen. Tidak seperti steroidogenesis, folikulogenesis bergantung pada FSH. Saat folikel tumbuh dan kadar estrogen meningkat, mekanisme umpan balik diaktifkan - produksi FSH dihambat, yang pada gilirannya menyebabkan penurunan aktivitas aromatase folikel dan, akhirnya, ke atresia folikel melalui apoptosis (kematian sel terprogram).
Mekanisme umpan balik estrogen dan FSH menghambat perkembangan folikel yang sudah mulai tumbuh, tetapi tidak pada folikel dominan. Folikel dominan mengandung lebih banyak reseptor FSH, yang mendukung proliferasi sel granulosa dan aromatisasi androgen menjadi estrogen. Selain itu, jalur parakrin dan autokrin berperan sebagai koordinator penting perkembangan folikel antral.
Pengatur autokrin/parakrin terdiri dari peptida (inhibin, aktivin, folistatin), yang disintesis oleh sel granulosa sebagai respons terhadap FSH dan memasuki cairan folikel. Inhibin mengurangi sekresi FSH; aktivin merangsang pelepasan FSH dari kelenjar pituitari dan meningkatkan aksi FSH di ovarium; folistatin menekan aktivitas FSH, mungkin dengan mengikat aktivin. Setelah ovulasi dan perkembangan korpus luteum, inhibin berada di bawah kendali LH.
Pertumbuhan dan diferensiasi sel ovarium dipengaruhi oleh faktor pertumbuhan mirip insulin (IGE). IGF-1 bekerja pada sel granulosa, menyebabkan peningkatan siklik adenosin monofosfat (cAMP), progesteron, oksitosin, proteoglikan, dan inhibin.
IGF-1 bekerja pada sel-sel teka, yang menyebabkan peningkatan produksi androgen. Sel-sel teka, pada gilirannya, menghasilkan faktor nekrosis tumor (TNF) dan faktor pertumbuhan epidermal (EGF), yang juga diatur oleh FSH.
EGF merangsang proliferasi sel granulosa. IGF-2 merupakan faktor pertumbuhan utama dalam cairan folikel, dan IGF-1, TNF-a, TNF-3, dan EGF juga ditemukan di dalamnya.
Gangguan regulasi parakrin dan/atau autokrin pada fungsi ovarium tampaknya berperan dalam gangguan ovulasi dan dalam perkembangan ovarium polikistik.
Seiring pertumbuhan folikel antral, kandungan estrogen dalam cairan folikel meningkat. Pada puncak peningkatannya, reseptor untuk LH muncul pada sel granulosa, terjadi luteinisasi sel granulosa, dan produksi progesteron meningkat. Jadi, pada periode praovulasi, peningkatan produksi estrogen menyebabkan munculnya reseptor LH, LH, pada gilirannya, menyebabkan luteinisasi sel granulosa dan produksi progesteron. Peningkatan progesteron mengurangi kadar estrogen, yang tampaknya menyebabkan puncak kedua FSH di tengah siklus.
Ovulasi diperkirakan terjadi 10-12 jam setelah puncak LH dan 24-36 jam setelah puncak estradiol. LH diperkirakan merangsang pembelahan reduksi oosit, luteinisasi sel granulosa, dan sintesis progesteron dan prostaglandin dalam folikel.
Progesteron meningkatkan aktivitas enzim proteolitik, yang bersama dengan prostaglandin berperan dalam pecahnya dinding folikel. Puncak FSH yang diinduksi progesteron memungkinkan oosit keluar dari folikel dengan mengubah plasminogen menjadi enzim proteolitik plasmin, dan menyediakan reseptor LH dalam jumlah yang cukup untuk perkembangan normal fase luteal.
Dalam waktu 3 hari setelah ovulasi, sel-sel granulosa bertambah besar, dan vakuola khas yang diisi dengan pigmen, lutein, muncul di dalamnya. Sel-sel teka-luteal berdiferensiasi dari teka dan stroma dan menjadi bagian dari korpus luteum. Kapiler yang menembus korpus luteum berkembang sangat cepat di bawah pengaruh faktor angiogenesis, dan dengan peningkatan vaskularisasi, produksi progesteron dan estrogen meningkat. Aktivitas steroidogenesis dan umur korpus luteum ditentukan oleh kadar LH. Korpus luteum bukanlah formasi seluler yang homogen. Selain 2 jenis sel luteal, ia mengandung sel endotel, makrofag, fibroblas, dll. Sel luteal besar menghasilkan peptida (relaksin, oksitosin) dan lebih aktif dalam steroidogenesis dengan aktivitas aromatase yang lebih besar dan sintesis progesteron yang lebih besar daripada sel-sel kecil.
Puncak progesteron terjadi pada hari ke-8 setelah puncak LH. Perlu dicatat bahwa progesteron dan estradiol disekresikan secara episodik pada fase luteal yang berkorelasi dengan produksi LH yang berdenyut. Dengan terbentuknya korpus luteum, kendali produksi inhibin berpindah dari FSH ke LH. Inhibin meningkat seiring dengan peningkatan estradiol sebelum puncak LH dan terus meningkat setelah puncak LH, meskipun kadar estrogen menurun. Meskipun inhibin dan estradiol disekresikan oleh sel granulosa, keduanya diatur oleh jalur yang berbeda. Penurunan inhibin pada akhir fase luteal berkontribusi pada peningkatan FSH untuk siklus berikutnya.
Korpus luteum menurun sangat cepat - pada hari ke 9-11 setelah ovulasi.
Mekanisme degenerasi tidak jelas dan tidak terkait dengan peran luteolitik estrogen atau mekanisme terkait reseptor, seperti yang diamati pada endometrium. Ada penjelasan lain untuk peran estrogen yang diproduksi oleh korpus luteum. Diketahui bahwa estrogen diperlukan untuk sintesis reseptor progesteron di endometrium. Estrogen fase luteal mungkin diperlukan untuk perubahan terkait progesteron di endometrium setelah ovulasi. Perkembangan reseptor progesteron yang tidak memadai, sebagai akibat dari kadar estrogen yang tidak memadai, mungkin merupakan mekanisme tambahan untuk infertilitas dan keguguran dini, bentuk lain dari defisiensi fase luteal. Dipercayai bahwa masa hidup korpus luteum terbentuk pada saat ovulasi. Dan itu pasti akan mengalami kemunduran jika tidak didukung oleh human chorionic gonadotropin karena kehamilan. Dengan demikian, regresi korpus luteum menyebabkan penurunan kadar estradiol, progesteron, dan inhibin. Penurunan inhibin menghilangkan efek supresifnya pada FSH; Penurunan estradiol dan progesteron memungkinkan pemulihan sekresi GnRH yang sangat cepat dan penghapusan mekanisme umpan balik dari kelenjar pituitari. Penurunan inhibin dan estradiol, bersama dengan peningkatan GnRH, menimbulkan FSH atas LH. Peningkatan FSH menyebabkan pertumbuhan folikel dengan pemilihan folikel dominan berikutnya, dan siklus baru dimulai jika kehamilan tidak terjadi. Hormon steroid memainkan peran utama dalam biologi reproduksi dan fisiologi umum. Mereka menentukan fenotipe manusia, memengaruhi sistem kardiovaskular, metabolisme tulang, kulit, kesejahteraan umum dan memainkan peran kunci dalam kehamilan. Tindakan hormon steroid mencerminkan mekanisme intraseluler dan genetik yang diperlukan untuk mengirimkan sinyal ekstraseluler ke inti sel untuk menyebabkan respons fisiologis.
Estrogen menembus membran sel secara difus dan mengikat reseptor yang terletak di inti sel. Kompleks reseptor-steroid kemudian mengikat DNA. Pada sel target, interaksi ini mengarah pada ekspresi gen, sintesis protein, dan fungsi sel dan jaringan tertentu.
Использованная литература