
Semua konten iLive ditinjau secara medis atau diperiksa fakta untuk memastikan akurasi faktual sebanyak mungkin.
Kami memiliki panduan sumber yang ketat dan hanya menautkan ke situs media terkemuka, lembaga penelitian akademik, dan, jika mungkin, studi yang ditinjau secara medis oleh rekan sejawat. Perhatikan bahwa angka dalam tanda kurung ([1], [2], dll.) Adalah tautan yang dapat diklik untuk studi ini.
Jika Anda merasa salah satu konten kami tidak akurat, ketinggalan zaman, atau dipertanyakan, pilih dan tekan Ctrl + Enter.
Penyebab dan patogenesis difteri
Ahli medis artikel

Terakhir ditinjau: 04.07.2025
Difteri merupakan paradigma penyakit menular yang bersifat toksigenik. Pada tahun 1883, Klebs menunjukkan bahwa Corynebacterium diphtheriae merupakan agen penyebab difteri. Setahun kemudian, Loeffler menemukan bahwa organisme tersebut hanya dapat dibiakkan dari rongga nasofaring dan mengusulkan bahwa kerusakan pada organ dalam disebabkan oleh toksin yang dapat larut. Pada tahun 1888, Roux dan Yersin menunjukkan bahwa hewan yang disuntik dengan filtrat steril C. diphtheriae mengembangkan patologi organ yang tidak dapat dibedakan dari difteri manusia; hal ini menunjukkan bahwa eksotoksin yang kuat merupakan faktor virulensi utama.
Difteri paling sering merupakan infeksi saluran pernapasan atas yang menyebabkan demam, sakit tenggorokan, dan malaise. Selaput fibrin tebal berwarna abu-abu kehijauan, pseudomembran, sering terbentuk di lokasi infeksi sebagai akibat dari efek gabungan pertumbuhan bakteri, produksi toksin, nekrosis jaringan di bawahnya, dan respons imun inang. Pengakuan bahwa kerusakan organ sistemik disebabkan oleh aksi toksin difteri telah mengarah pada pengembangan terapi berbasis antitoksin yang efektif untuk pengobatan infeksi akut dan vaksin toksoid yang sangat efektif.
Meskipun imunisasi toksoid telah menjadikan difteri sebagai penyakit langka di daerah-daerah yang standar kesehatan masyarakatnya mengharuskan vaksinasi, wabah difteri masih terjadi pada kelompok yang tidak diimunisasi dan yang kekebalan tubuhnya lemah. Sebaliknya, wabah difteri yang meluas dan mencapai proporsi epidemik telah terjadi di daerah-daerah yang program imunisasinya telah dihentikan.
Jenis corynebacteria lainnya
Selain C. diphtheriae, C. ulcerans, dan C. pseudotuberculosis, C. pseudodiphtheriticum dan C. xerosis terkadang dapat menyebabkan infeksi nasofaring dan kulit. Dua galur terakhir dapat dikenali dari kemampuannya menghasilkan pirazinamidase. Dalam kedokteran hewan, C. renale dan C. kutscheri merupakan patogen penting, yang masing-masing menyebabkan pielonefritis pada sapi dan infeksi laten pada tikus.
Penyebab
Agen penyebab difteri, Corynebacterium diphtheriae, merupakan batang tipis, sedikit melengkung dengan penebalan berbentuk tongkat di ujungnya, tidak bergerak; tidak membentuk spora, kapsul atau flagela, dan bersifat gram positif.
Selain toksin, corynebacteria difteri menghasilkan neuraminidase, hyaluronidase, hemolisin, faktor nekrosis dan faktor difus selama proses hidupnya, yang dapat menyebabkan nekrosis dan pencairan substansi utama jaringan ikat.
Berdasarkan kemampuannya membentuk toksin, korinebakteria difteri dibagi menjadi toksigenik dan non-toksigenik.
Toksin difteri adalah eksotoksin bakteri yang kuat yang menentukan manifestasi klinis umum dan lokal dari penyakit ini. Toksisitas ditentukan secara genetik. Corynebacteria difteri yang tidak bersifat toksigenik tidak menyebabkan penyakit ini.
Berdasarkan ciri-ciri kultural dan morfologis, semua corynebacteria difteri dibagi menjadi 3 varian: gravis, mitis, intermedius. Tidak ada ketergantungan langsung tingkat keparahan penyakit pada varian corynebacteria difteri. Setiap varian mengandung strain toksigenik dan non-toksigenik. Corynebacteria toksigenik dari semua varian menghasilkan toksin yang identik. [ 1 ]
Struktur, Klasifikasi dan Jenis Antigen
Corynebacterium diphtheriae adalah bakteri Gram-positif, batang non-motil, berbentuk gada. Strain yang tumbuh dalam jaringan atau kultur in vitro yang lebih tua mengandung bintik-bintik halus di dinding sel yang memungkinkan terjadinya dekolorisasi selama pewarnaan Gram dan menghasilkan reaksi Gram yang bervariasi. Kultur yang lebih tua sering mengandung butiran metakromatik (polimetafosfat) yang diwarnai ungu kebiruan dengan biru metilen. Gula dinding sel meliputi arabinosa, galaktosa, dan manosa. Selain itu, ester trehalosa 6,6' yang beracun dapat diisolasi, yang mengandung asam korinemikolat dan korinemikolenat dalam konsentrasi yang sama. Tiga jenis kultur yang berbeda dikenali: mitis, intermedius, gravis.
Sebagian besar strain membutuhkan asam nikotinat dan pantotenat untuk pertumbuhan; beberapa juga membutuhkan tiamin, biotin, atau asam pimelat. Untuk produksi toksin difteri yang optimal, medium harus dilengkapi dengan asam amino dan harus disisihkan.
Sejak awal tahun 1887, Loeffler menggambarkan isolasi C. diphtheriae yang avirulen (nontoksigenik) yang tidak dapat dibedakan dari galur virulen (toksigenik) yang diisolasi dari pasien pada individu yang sehat. Sekarang diketahui bahwa galur C. diphtheriae yang avirulen dapat diubah menjadi fenotipe virulen setelah infeksi dan lisogenisasi oleh salah satu dari sejumlah korinebakteriofaga yang membawa gen struktural untuk toksin difteri, tox. Konversi lisogenik dari fenotipe avirulen menjadi virulen dapat terjadi baik secara in situ maupun in vitro. Gen struktural untuk toksin difteri tidak penting bagi korinebakteriofaga maupun C. diphtheriae. Meskipun ada pengamatan ini, pergeseran genetik toksin difteri belum diamati.
 [
[ Patogenesis
Titik masuk infeksi adalah selaput lendir orofaring, hidung, laring, lebih jarang selaput lendir mata dan alat kelamin, serta kulit yang rusak, permukaan luka atau luka bakar, ruam popok, luka pusar yang belum sembuh. Di titik masuk tersebut, bakteri corynebacterium difteri berkembang biak dan mengeluarkan eksotoksin.
Eksudat yang kaya akan fibrinogen dikeluarkan dan diubah menjadi fibrin di bawah pengaruh trombokinase yang dilepaskan selama nekrosis sel epitel. Lapisan fibrin terbentuk - tanda khas difteri.
Pengangkutan nasofaring asimtomatik umum terjadi di daerah endemis difteri. Pada individu yang rentan, galur toksigenik menyebabkan penyakit dengan mereplikasi dan mengeluarkan toksin difteri di nasofaring atau lesi kulit. Lesi difteri sering kali ditutupi oleh pseudomembran yang terdiri dari fibrin, bakteri, dan sel inflamasi. Toksin difteri dapat dibelah secara proteolitik menjadi dua fragmen: fragmen N-terminal A (domain katalitik) dan fragmen B (domain transmembran dan pengikat reseptor). Fragmen A mengkatalisis ribosilasi ADP yang bergantung pada NAD+ dari faktor pemanjangan 2, sehingga menghambat sintesis protein dalam sel eukariotik. Fragmen B mengikat reseptor permukaan sel dan memfasilitasi pengiriman fragmen A ke sitosol.
Kekebalan protektif melibatkan respons antibodi terhadap toksin difteri setelah penyakit klinis atau terhadap toksin difteri (toksin yang dinonaktifkan oleh formaldehida) setelah imunisasi.
Kolonisasi
Sedikit yang diketahui tentang faktor-faktor yang memediasi kolonisasi C. diphtheriae. Akan tetapi, jelas bahwa faktor-faktor selain produksi toksin difteri berkontribusi terhadap virulensi. Studi epidemiologi telah menunjukkan bahwa lisotipe tertentu dapat bertahan dalam suatu populasi untuk jangka waktu yang lama. Lisotipe tersebut kemudian dapat digantikan oleh lisotipe lain. Kemunculan dan dominasi selanjutnya dari lisotipe baru dalam suatu populasi diduga terkait dengan kemampuannya untuk berkoloni dan bersaing secara efektif dalam segmen ceruk ekologi nasofaringnya. Corynebacterium diphtheriae dapat menghasilkan neuraminidase, yang memecah asam sialik permukaan sel menjadi komponen-komponennya, piruvat dan asam N-asetilneuraminat. Faktor korda (6,6'-di-O-mikoloil-α, α'-D-trehalosa) merupakan komponen permukaan C. diphtheriae, tetapi perannya dalam kolonisasi inang manusia tidak jelas.
Produksi toksin difteri
Gen struktural toksin difteri, tox, termasuk dalam famili corynebacteriophage yang berkerabat dekat, di antaranya β-phage adalah yang paling banyak dipelajari. Pengaturan ekspresi toksin difteri dimediasi oleh represor yang diaktifkan oleh zat besi, DtxR, yang dikodekan oleh genom C. diphtheriae. Ekspresi toksin bergantung pada keadaan fisiologis C. diphtheriae. Dalam kondisi di mana zat besi menjadi substrat pembatas laju pertumbuhan, zat besi terdisosiasi dari DtxR, gen toxicodendron menjadi aktif, dan toksin difteri disintesis dan disekresikan ke dalam media kultur pada laju maksimal.
Toksin difteri sangat kuat; untuk spesies yang rentan (misalnya manusia, monyet, kelinci, marmut), dosis 100 hingga 150 ng/kg berat badan sudah mematikan. Toksin difteri terdiri dari rantai polipeptida tunggal yang terdiri dari 535 asam amino. Analisis biokimia, genetik, dan struktur sinar-X menunjukkan bahwa toksin tersebut terdiri dari tiga domain struktural/fungsional:
- ADP-ribosiltransferase N-terminal (domain katalitik);
- wilayah yang memfasilitasi pengiriman domain katalitik melintasi membran sel (domain transmembran);
- domain pengikatan reseptor sel eukariotik.
Setelah pencernaan tripsin ringan dan reduksi dalam kondisi denaturasi, toksin difteri dapat dibelah secara spesifik pada loop peka protease menjadi dua fragmen polipeptida (A dan B). Fragmen A adalah komponen N-terminal 21 kDa dari toksin dan mengandung situs katalitik untuk ADP-ribosilasi faktor elongasi 2 (EF-2)
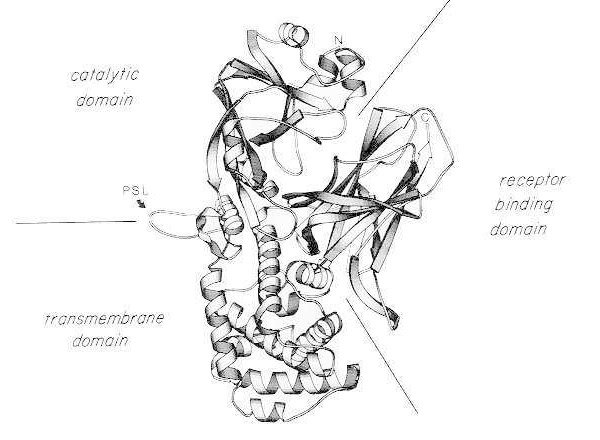
Diagram pita struktur kristal sinar-X dari toksin difteri asli monomerik. (dimodifikasi dari Bennett MJ, Choe S, Eisenberg D: Domain swapping: Entangling aliances between protein. Proc Natl Acad Sci, USA, 91: 3127, 1994). Posisi relatif dari domain katalitik, transmembran, dan pengikat reseptor ditunjukkan. Toksin utuh dapat dibelah oleh protease mirip tripsin di Arg190, Arg192, dan/atau Arg193, yang terletak di loop sensitif protease (PSL). Setelah reduksi jembatan disulfida antara Cys186 dan Cys201, toksin dapat diurai menjadi fragmen A dan B. Ujung amino (N) dan ujung karboksi (C) dari toksin utuh ditunjukkan. Bagan strip dibuat menggunakan program MOLESCRIPT.
Fragmen C-terminal, fragmen B, membawa domain transmembran dan pengikatan reseptor toksin.
Keracunan sel eukariotik tunggal oleh toksin difteri melibatkan setidaknya empat langkah berbeda:
- pengikatan toksin ke reseptornya di permukaan sel;
- pengelompokan reseptor bermuatan ke dalam lubang berlapis dan internalisasi toksin melalui endositosis yang dimediasi reseptor; setelah pengasaman vesikel endositosis oleh pompa proton yang digerakkan oleh ATP yang terkait dengan membran,
- penyisipan domain transmembran ke dalam membran dan memfasilitasi pengiriman domain katalitik ke sitosol, dan
- ADP-ribosilasi EF-2, mengakibatkan penekanan sintesis protein yang ireversibel.
Telah ditunjukkan bahwa satu molekul domain katalitik yang dikirim ke sitosol cukup untuk mematikan sel.
Diagram skema keracunan difteri pada sel eukariotik yang rentan.
Toksin tersebut mengikat reseptor permukaan selnya dan diinternalisasi melalui endositosis yang dimediasi reseptor; setelah pengasaman endosom, domain transmembran dimasukkan ke dalam membran vesikel; domain katalitik dikirimkan ke sitosol, yang menyebabkan penghambatan sintesis protein dan kematian sel.
Epidemiologi
Sebelum imunisasi massal penduduk AS dengan toksoid difteri, difteri biasanya merupakan penyakit anak-anak. Aspek yang luar biasa dari imunisasi massal dengan toksoid difteri adalah bahwa seiring dengan meningkatnya persentase penduduk dengan tingkat kekebalan antitoksin yang protektif (≥ 0,01 IU/mL), frekuensi isolasi galur toksigenik dari penduduk menurun. Saat ini, di Amerika Serikat, di mana difteri klinis hampir sepenuhnya menghilang, isolasi galur toksigenik C. diphtheriae jarang terjadi. Karena infeksi subklinis tidak lagi menyediakan sumber paparan antigen difteri dan, kecuali ditingkatkan, kekebalan terhadap antitoksin berkurang, sebagian besar orang dewasa (30 hingga 60%) memiliki kadar antitoksin di bawah tingkat protektif dan berisiko. Di Amerika Serikat, Eropa, dan Eropa Timur, wabah difteri baru-baru ini terjadi terutama di antara individu yang menyalahgunakan alkohol dan/atau narkoba. Dalam kelompok ini, pembawa C. diphtheriae toksigenik memiliki tingkat kekebalan antitoksik yang cukup tinggi. Gagalnya langkah-langkah kesehatan masyarakat di Rusia baru-baru ini telah mengakibatkan difteri menjadi epidemi. Pada akhir tahun 1994, lebih dari 80.000 kasus dan lebih dari 2.000 kematian telah dilaporkan di Rusia.
Wabah fokal difteri hampir selalu dikaitkan dengan pembawa kekebalan yang kembali dari wilayah endemik difteri. Bahkan, wabah difteri klinis baru-baru ini di Amerika Serikat dan Eropa dikaitkan dengan pelancong yang kembali dari Rusia dan Eropa Timur. Strain toksigenik C. diphtheriae ditularkan langsung dari orang ke orang melalui droplet di udara. Diketahui bahwa strain toksigenik dapat langsung menjajah rongga nasofaring. Selain itu, gen toksigenik dapat disebarkan secara tidak langsung melalui pelepasan korinebakteriofag toksigenik dan transformasi lisogenik C. diphtheriae autokton nontoksigenik in situ. [ 5 ]
Selain penentuan biotipe dan lisotipe isolat C. diphtheriae, teknik biologi molekuler kini dapat digunakan untuk mempelajari wabah difteri. Pola pencernaan endonuklease restriksi DNA kromosom C. diphtheriae telah digunakan untuk mempelajari wabah klinis, seperti halnya penggunaan sekuens penyisipan korinebakteri kloning sebagai pemeriksaan genetik.
Uji Schick telah digunakan selama bertahun-tahun untuk menilai kekebalan terhadap toksin difteri, meskipun sekarang telah digantikan di banyak daerah oleh uji serologis untuk antibodi spesifik terhadap toksin difteri. Dalam uji Schick, sejumlah kecil toksin difteri (sekitar 0,8 ng dalam 0,2 ml) disuntikkan secara intradermal ke lengan bawah (lokasi uji) dan 0,0124 μg toksin difteri dalam 0,2 ml disuntikkan secara intradermal ke lokasi kontrol. Pembacaan dilakukan pada 48 dan 96 jam. Reaksi kulit nonspesifik biasanya mencapai puncaknya pada 48 jam. Pada 96 jam, reaksi eritematosa dengan beberapa kemungkinan nekrosis di lokasi uji menunjukkan kekebalan antitoksik yang tidak cukup untuk menetralkan toksin (≤ 0,03 IU/ml). Peradangan di area uji dan kontrol setelah 48 jam menunjukkan reaksi hipersensitivitas terhadap sediaan antigen.
Formulir
Corynebacterium diphtheriae menginfeksi nasofaring atau kulit. Strain toksigenik menghasilkan eksotoksin kuat yang dapat menyebabkan difteri. Gejala difteri meliputi faringitis, demam, dan pembengkakan pada leher atau area di sekitar lesi kulit. Lesi difteri ditutupi oleh pseudomembran. Toksin menyebar melalui aliran darah ke organ-organ yang jauh dan dapat menyebabkan kelumpuhan dan gagal jantung kongestif. [ 6 ]
Terdapat dua jenis difteri klinis: nasofaring dan kutan. Gejala difteri faring berkisar dari faringitis ringan hingga hipoksia akibat obstruksi jalan napas oleh pseudomembran. Keterlibatan kelenjar getah bening serviks dapat menyebabkan pembengkakan parah pada leher (difteri leher banteng), dan pasien dapat mengalami demam (≥ 103°F). Lesi kulit pada difteri kutan biasanya ditutupi oleh pseudomembran abu-abu kecokelatan. Komplikasi sistemik yang mengancam jiwa, terutama hilangnya fungsi motorik (misalnya, kesulitan menelan) dan gagal jantung kongestif, dapat berkembang sebagai akibat dari aksi toksin difteri pada neuron motorik perifer dan miokardium.
Kontrol
Pengendalian difteri bergantung pada imunisasi yang memadai dengan toksoid difteri: toksin difteri yang dinonaktifkan oleh formaldehida, yang tetap utuh secara antigenik. Toksoid disiapkan dengan menginkubasi toksin difteri dengan formaldehida pada suhu 37°C dalam kondisi basa. Imunisasi difteri harus dimulai pada bulan kedua kehidupan dengan serangkaian tiga dosis primer dengan interval 4 hingga 8 minggu, diikuti dengan dosis keempat sekitar 1 tahun setelah dosis primer terakhir. Toksoid difteri banyak digunakan sebagai komponen vaksin difteri-pertusis-tetanus (DPT). Studi epidemiologi telah menunjukkan bahwa imunisasi terhadap difteri sekitar 97% efektif. Meskipun imunisasi massal terhadap difteri dipraktikkan di Amerika Serikat dan Eropa dan tingkat imunisasi anak-anak memadai, sebagian besar populasi orang dewasa mungkin memiliki titer antibodi di bawah tingkat protektif. Orang dewasa harus divaksinasi ulang dengan toksoid difteri setiap 10 tahun. Memang, imunisasi penguat dengan toksoid difteri-tetanus harus diberikan kepada pelancong ke daerah dengan tingkat difteri endemik yang tinggi (Amerika Tengah dan Selatan, Afrika, Asia, Rusia, dan Eropa Timur). Dalam beberapa tahun terakhir, penggunaan sediaan toksoid yang sangat murni untuk imunisasi telah meminimalkan reaksi hipersensitivitas berat yang terjadi sesekali. imunisasi penguat dengan toksoid difteri-tetanus harus diberikan kepada pelancong ke daerah dengan tingkat difteri endemik yang tinggi (Amerika Tengah dan Selatan, Afrika, Asia, Rusia, dan Eropa Timur). Dalam beberapa tahun terakhir, penggunaan sediaan toksoid yang sangat murni untuk imunisasi telah meminimalkan reaksi hipersensitivitas berat yang terjadi sesekali. Imunisasi penguat dengan toksoid difteri-tetanus harus diberikan kepada pelancong ke daerah dengan tingkat difteri endemik yang tinggi (Amerika Tengah dan Selatan, Afrika, Asia, Rusia, dan Eropa Timur). Dalam beberapa tahun terakhir, penggunaan sediaan toksoid yang sangat murni untuk imunisasi telah meminimalkan reaksi hipersensitivitas berat yang terjadi sesekali.
Meskipun antibiotik (seperti penisilin dan eritromisin) digunakan sebagai bagian dari pengobatan pasien difteri, imunisasi pasif cepat dengan antitoksin difteri paling efektif dalam mengurangi kematian kasus. Waktu paruh antitoksin spesifik yang panjang dalam aliran darah merupakan faktor penting dalam memastikan netralisasi toksin difteri yang efektif; namun, agar efektif, antitoksin harus bereaksi dengan toksin sebelum dapat memasuki sel.
Rekayasa ulang toksin difteri untuk pengembangan sitotoksin spesifik reseptor eukariotik
Rekayasa protein merupakan bidang baru dan berkembang pesat dalam biologi molekuler; bidang ini menggabungkan metodologi DNA rekombinan dan sintesis DNA fase padat untuk merancang dan membangun gen kimerik yang produknya memiliki sifat unik. Studi tentang hubungan struktur-fungsi toksin difteri telah menunjukkan dengan jelas bahwa toksin ini merupakan protein tiga domain: katalitik, transmembran, dan reseptor. Telah dimungkinkan untuk mengganti secara genetik domain pengikatan reseptor asli toksin difteri dengan berbagai hormon polipeptida dan sitokin (misalnya, hormon perangsang α-melanosit [α-MSH], interleukin (IL) 2, IL-4, IL-6, IL-7, faktor pertumbuhan epidermal). Protein kimerik atau toksin fusi yang dihasilkan menggabungkan spesifisitas pengikatan reseptor sitokin dengan domain transmembran dan katalitik toksin. Dalam setiap kasus, toksin fusi terbukti secara selektif hanya meracuni sel-sel yang memiliki reseptor target yang sesuai. Toksin fusi yang dimodifikasi secara genetik pertama ini, DAB 389IL-2, saat ini sedang dievaluasi dalam uji klinis pada manusia untuk pengobatan limfoma refrakter dan penyakit autoimun di mana sel dengan reseptor IL-2 afinitas tinggi memainkan peran penting dalam patogenesis.[ 7 ] Pemberian DAB 389 IL-2 terbukti aman, ditoleransi dengan baik, dan mampu menginduksi remisi penyakit yang tahan lama tanpa efek samping yang serius. Toksin fusi berbasis toksin difteri kemungkinan akan menjadi agen biologis baru yang penting untuk pengobatan tumor atau penyakit tertentu di mana reseptor permukaan sel tertentu dapat ditargetkan.